
NIVEL DE OXIGENACIÓN DEL AGUA
Además de la condición estacional de la temperatura, ésta influye en las comunidades a partir de una propiedad innata que tiene por su naturaleza química, y es que cuando está fría es capaz de disolver mayor cantidad de oxígeno que cuando está más caliente.
Benson & Krause (1980) midieron la solubilidad del gas oxígeno para el agua pura en un rango desde 16,6 mg/L cuando está a 0 ºC , hasta 6,4 mg/L para 40 ºC, [en condiciones de presión ambiental normal y aire saturado de vapor].
Esto es de gran relevancia en la ecología de las masas de agua para los animales y otros seres vivos que necesitan respirar del oxígeno que está disuelto en el agua.
Un rango deseable de oxígeno disuelto (10 – 8 mg/L) para soportar la vida animal en el interior de la masa de agua por lo tanto es posible en condiciones de temperaturas de 16-25 ºC, habituales hasta ahora para los lagos ibéricos en verano. Sin embargo con el incremento de las temperaturas registrado en los últimos años en toda Europa se están sobrepasando los límites importantes para las comunidades bióticas.
Además la temperatura también influye en el nivel de disolución de otras sustancias, por lo que es especialmente relevante también en la monitorización de otros dos parámetros importantes en limnología: el pH y la conductividad.
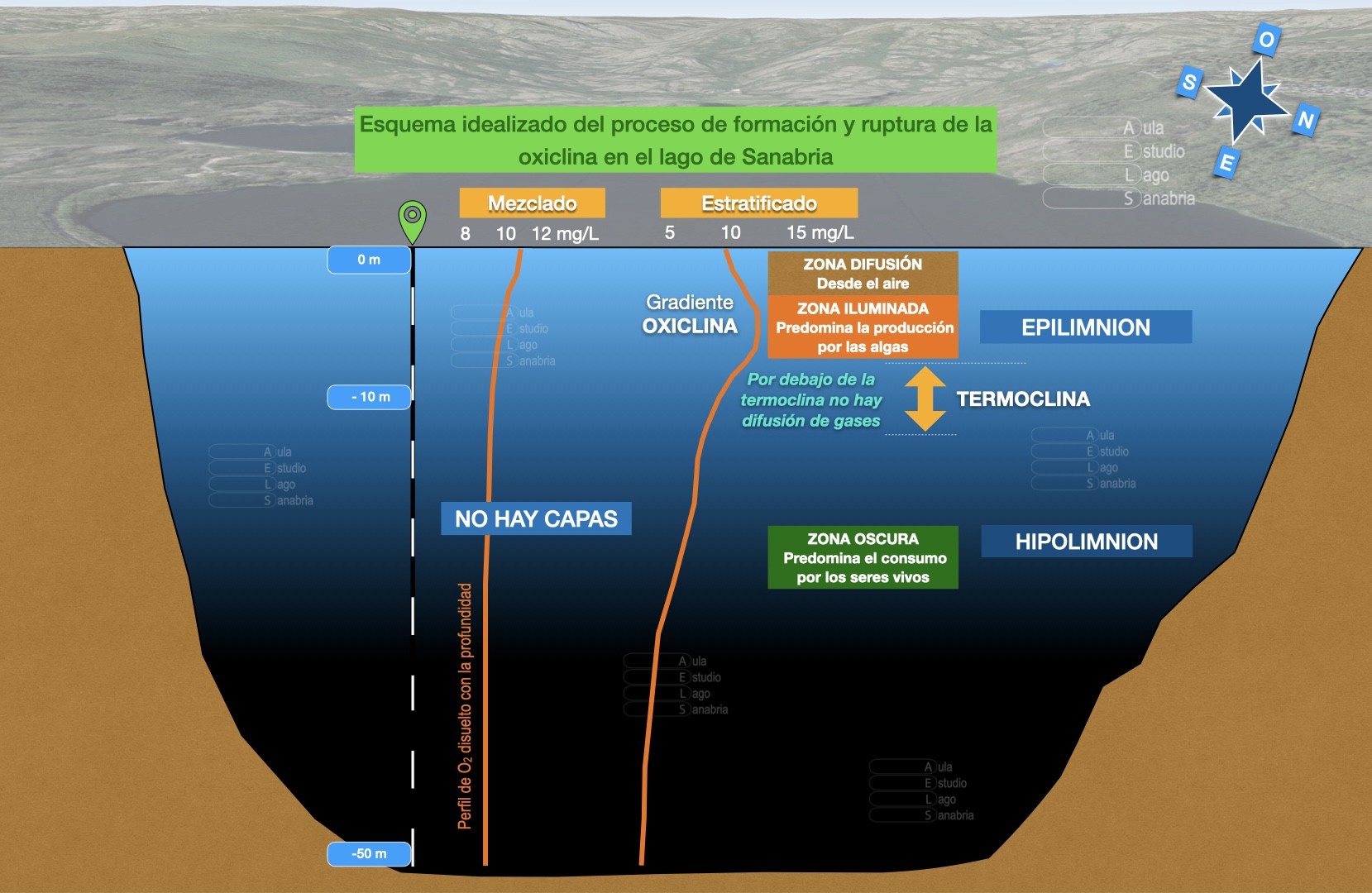
La incidencia que tiene la temperatura a lo largo de la columna de agua con formación de capas de distinta densidad, controla a su vez la difusión de gases. De esta forma en la zona más superficial es posible la difusión de oxígeno desde el aire, mientras que en profundidad no es posible. Además dentro del agua la fotosíntesis producida por la algas es otra fuente endógena de oxígeno en el agua, pero limitada hacia abajo por la termoclina y por la presencia de luz.
Por lo tanto en las capas superficiales, por encima de la termoclina, predomina la difusión y la producción y aunque los seres vivos que allí viven consumen al respirar algo de oxígeno el balance es positivo. Se produce un gradiente llamado oxiclina. Por debajo de la temoclina el agua está más fría y es más densa, y además la luz se extingue. Cuanto más profundo menos posible es realizar fotosíntesis, de manera que en estas capas más profundas del hipolimnion predomina el consumo del oxígeno. Y además permanecen aisladas de las superficiales hasta que se rompa la estratificación y con la mezcla invernal llega a las zonas profundas agua con oxígeno disuelto.
Cuando la estratificación del fondo es muy larga (más tiempo de incomunicación con la superficie) y/o cuando hay mucho consumo biótico o químico del oxígeno disuelto que tiene el agua (respiración y oxidación química y microbiológica de la materia orgánica) se puede llegar a agotar. Por eso es muy relevante la reserva que tienen las capas profundas. En el caso de consumirse la reserva el fondo de la masa de agua sufre un proceso de hipoxia, y en el caso de reducción extrema o carencia total de oxígeno un proceso de anoxia.
Las mayores concentraciones de oxígeno disuelto se observan en la superficie del lago y en periodos de aguas frías, siguiendo la dinámica esperada; con máximos que superan 10,5 mg/L. Durante la fase de estratificación el contenido disminuye con el tiempo (por consumo interno) y con la profundidad (por falta de difusión desde la superficie); alcanzándose mínimos temporalmente en torno a 1,5 – 3 mg/L. Lo que supone niveles de hipoxia esperables a 50 m de profundidad tras unos cuantos meses de estancamiento del agua.
El gradiente de oxígeno disuelto presenta una forma curva típica para los lagos oligotróficos, con una oxiclina con pendiente ligeramente positiva con la profundidad en el epilimnion hasta llegar a la termoclina (en función de la disposición de las poblaciones de los seres fotosintéticos), y que después a lo largo de toda la columna del hipolimnion tiene una pendiente negativa (es la zona oscura, sin posibilidad de hacer fotosíntesis). Por lo tanto decrece de forma paulatina hasta casi el fondo, donde se alcanzan los mínimos absolutos; y donde dominan además procesos químicos reductores derivados de la actividad microbiana.
No se conocen episodios de anoxia en el fondo del lago (el hipolimnion) a lo largo de una serie de años muy larga con medidas mensuales en contínuo, de manera que por el momento la regeneración de toda la masa de agua en la fase de mezcla es completa. Por lo tanto el sistema es capaz de “digerir” toda la carga orgánica que llega por la cuenca y la producción endógena por crecimiento algal, y que también sedimenta hacia las aguas profundas, con el oxígeno que almacena en profundidad desde cuando se forma la estratificación hasta que se inicia la mezcla.

Gráficas creadas con los valores disponibles en la web de la Fundación Patrimonio Natural de CyL


Gráficas creada con los valores disponibles en la web de la Fundación Patrimonio Natural de CyL

A lo largo de la sucesión de los meses en una anualidad de calendario se producen variaciones en las concentraciones de este gas disuelto, y que siguen una misma pauta en todas las profundidades de la masa de agua.
Si analizamos los valores en la superficie (donde domina la difusión desde el aire) observamos máximos de finales de invierno (superiores a 10 mg/L en muchos años), y mínimos que coinciden el periodo más cálido del estiaje. En una relación directa e inversa con la temperatura del aire y el nivel de radiación solar sobre la superficie del lago. En este gráfico podemos observar un estrecho rango de datos, habitualmente comprendido entre 8,5 y 10,5 mg/L.
En la zona más profunda del epilimnion (hasta unos 8 a 10 m) se produce un ritmo de evolución del contenido de oxígeno disuelto simétrico al descrito en la superficie; pero para un rango de datos absolutos incluso mayores. Esto es debido a la liberación del gas por parte de las algas en todo el periodo de crecimiento vegetativo; y por lo tanto los máximos son de mediados o finales de primavera. Siendo los mínimos también de finales del verano, cuando se ralentiza o directamente cesa la producción fotosintética y la temperatura del agua en superficie impide la difusión hacia la columna de la masa acuática. En este segundo nivel de la columna los rangos habituales cada año oscilan más, entre 6,5 y 12 mg/L.
El mismo patrón encontramos en las profundidades, ya que entre otras cosas se produce una mezcla vertical del agua durante muchos meses. Los máximos (de hasta casi 10 mg/L) se alcanzan al final del periodo de la mezcla, en primavera; y los mínimos después de terminar la producción primaria en aguas de la zona iluminada y recibir una buena parte de la materia orgánica que allí se ha producido durante el verano. Y justo antes de comenzar la mezcla invernal que recargará de nuevo oxígeno disuelto en las aguas profundas. En este caso el rango de datos registrados en este lago es habitualmente de entre 10 y 3 mg/L, pero con mínimos puntuales más bajos como se ha indicado en otros apartados de esta entrada.
Estacionalmente se aprecia una evolución similar a lo largo de años de estudio. En el invierno, con el agua más fría y más agitada en superficie por el oleaje, se producen los niveles más elevados de oxigenación. Lo que induce una mayor variación interanual. En primavera y otoño se producen rangos de datos muy similares, y alternativos según el año. Sin encontrar un patrón de regularidad, dada la dependencia de la climatología de cada año en esas épocas (cada vez más inestable debido a las oscilaciones climáticas inducidas por efecto del cambio climático global).
Siendo en el verano cuando, incluso en superficie, se han registrado los valores promedio menores de toda la serie de datos. Muy constantes en torno a 7,9-8,3 mg/L.
Fuente: valores disponibles en la web de Fundación Patrimonio Natural de CyL
Puedes leer más sobre esto en:
- Hutchinson, G.E. 1957. A Treatise of Limnology. New York John Wiley & Sons.
- Wetzel, R.G. 1981. Limnología (Cap. 5). Ed. Omega.
- Wetzel, R.G & Likens G. 1991. Limonological Analyses. Second Edition. Ed. Springer-Verlag.
- Wetzel, R.G. 2001. Limnology (Chap. 5). Lake and River ecosystems. 3ª Ed. Elsevier.
- Margalef R. 1983. Limnología. Ed. Omega.
- De Hoyos C. 1996. Limnología del lago de Sanabria. Variabilidad interanual del fitoplancton. Tesis Doctoral. Universidad de Salamanca.
- Dodds W.K. 2002. Freshwater Ecology. Concepts and Environmental Applications. Ed. Academic Press & Elsevier Science Imprint. San Diego EEUU. 351 pp.


{kind=link}